深圳华大生命科学研究院、爱丁堡大学、约翰•霍普金斯大学、纽约大学、清华大学、天津大学等多家单位,通过开发酿酒酵母基因组序列设计与染色体构建技术,从头设计与全合成了酿酒酵母II号染色体(synII,长770 Kb),并成功导入酵母细胞,合成酵母菌株展现出与野生型高度相似的生命活性。此次国际协作组对真核生物酿酒酵母基因组的成功改造,是继人工支原体后人工生命创造领域又一里程碑式进展。该成果在2017年3月10日以封面专刊文章的形式发表于国际顶级学术期刊Science,入选2017年度中国科学十大进展,以下是共同第一作者王云博士详细解读。

文章题目:Deep functional analysis of synII, a 770-kilobase synthetic yeast chromosome
发表时间:2017-03-10
发表期刊:Science
主要研究团队:深圳华大生命科学研究院、爱丁堡大学、约翰•霍普金斯大学、纽约大学、清华大学、天津大学
影响因子:63.714
DOI:10.1126/science.aaf4791
研究背景
酵母是与人类生活关系最密切的一类单细胞真核微生物,在啤酒、面包的制作中不可或缺。自然界分布有超过1500种酵母,其中酿酒酵母(Saccharomyces cerevisiae)是最早被广泛用于食品发酵、疫苗生产等现代工业的酵母菌,是名副其实的“细胞工厂”;同时因为其清晰的遗传背景,成为科学研究最常用的模式生物之一。早在1996年,完成了酿酒酵母全基因组测序,并发现酵母基因组总共约6000个基因中有5000个不是维持生命活动所必需的,可以进行删减和改写。随着合成DNA技术的快速发展,大尺度重编基因组与人工合成成为可能。近几年,科学家通过非生命的化学物质组装成染色体,研究导致细胞死亡、细胞失活、生长缺陷的各项关键要素,探索生命本质。2010年,Gibson等人合成了首个原核生命体(生殖道支原体,Mycoplasma mycoides),而更复杂的真核基因组的序列设计与合成,已成为国际上合成基因组领域亟待突破的技术瓶颈。为此,研究团队人工设计并再造了酿酒酵母II号染色体,并深入解析了合成染色体生物学功能。
研究样本
野生型酿酒酵母(BY4741和BY4742);人工构建的一系列合成型酿酒酵母菌株;染色体着丝粒(CEN2) GFP荧光标记菌株。
以上菌株均由深圳国家基因库样本库储存。
研究策略
通过计算机辅助设计,重编酿酒酵母II号染色体序列,再通过从头化学合成DNA,逐级组装构建完整的合成染色体,通过监测细胞生长、DNA复制、细胞分裂,以及贯穿组学(Trans-Omics)分析等,深入解析合成酿酒酵母II号染色体的生物学功能,探索酵母人工染色体序列设计的理论准则。
研究成果
酵母人工染色体序列设计
研究团队首先利用合成酿酒酵母专用的计算机辅助序列设计工具BioStudio(约翰•霍普金斯大学,同期 Science专刊文章),遵循“强健、稳定、可变”三原则对酿酒酵母II号染色体进行了系统地序列设计。合成酵母II号染色体(chr02_3.25),总长为 770,035 bp,每467 bp发生了一次碱基设计更改,共发生了~10.26%的序列改变。与野生染色体相比,删除了22个introns、13个tRNA基因和8个转座子;替换90个TAG为 TAA,左右端粒被替换;通过同义密码子替换设计了532对28bp的PCRTag,并插入267个loxPsym位点(图1)。
在序列设计上,合成型酵母基因组最显著的特征在于引入了大量loxPsym特异重组位点。该位点可被一种可诱导产生的特异重组酶(Cre)识别并引发合成染色体重组(简称SCRaMbLE)。与野生单向重组loxP位点相比,loxPsym位点(34 bp序列:ATAACTTCGTATAATGTACATTATACGAAGTTAT)具有反向回文对称序列结构,使其具有诱发双向连续DNA重组的特性与能力 (SCRaMbLE,2016年 Genome Research 文章)。

图1. 酵母人工染色体序列设计
酵母人工染色体快速构建
为了快速合成与构建人工设计的染色体,研究团队将合成染色体序列分级拆分成不同大小的DNA合成片段(图2A)。利用优化的一步法Gibson组装将合成基因(Minichunk,~3 kb)组装成DNA长片段(ChunK ~10 kb)(图2B);采用SWAP-IN策略,同时转化3-5个长片段DNA至酵母细胞内,通过同源重组替换染色体中对应野生区(~ 30 kb)(图2C);最后,经过左臂18轮与右臂8轮的并行迭代替换与I-SceI介导的染色体整合,实现了完整的合成染色体人工构建(图2D)。本研究开发了I-SceI介导的染色体整合技术,将半合成染色体发生整合的效率可提高了10倍,同时合成染色体并行构建,显著提高了长染色体精准构建的效率,缩短时间近50%。

图2. 酵母人工染色体快速构建
人工合成染色体结构变异快速修复
酿酒酵母全合成染色体菌株 (chr02_9.01),经过高通量基因组测序,发现2个多拷贝重复的结构变异(CNV),分别位于megachunk L和T,重复区段总长分别为~15 kb与30 kb(图3B)。为了修复合成染色体中可能出现的高拷贝重复变异,利用I-SceI 蛋白识别并切割其特异位点,产生染色体双链断裂(DSB)诱发序列同源重组的原理,建立了一套简易的、高效的无标记多拷贝串联重复序列修复方法(图3A)。
首先,将带有URA3标记基因的同源插入整合片段和带有I-SceI蛋白表达盒的载体共转化,并在尿嘧啶(Ura)与组氨酸(His)营养缺陷培养基中筛选阳性克隆,最后利用半乳糖诱导I-SceI蛋白表达,引起合成染色体在I-SceI位点产生双链断裂,诱发同源重组,并用5-氟乳清酸(FOA)药物筛选无标记修复菌株。通过上述方法,完成合成染色体中的高拷贝重复变异修复,并经高通量基因组测序分析,最终获得正确的酿酒酵母菌株(chr02_9.03)(图3C)。
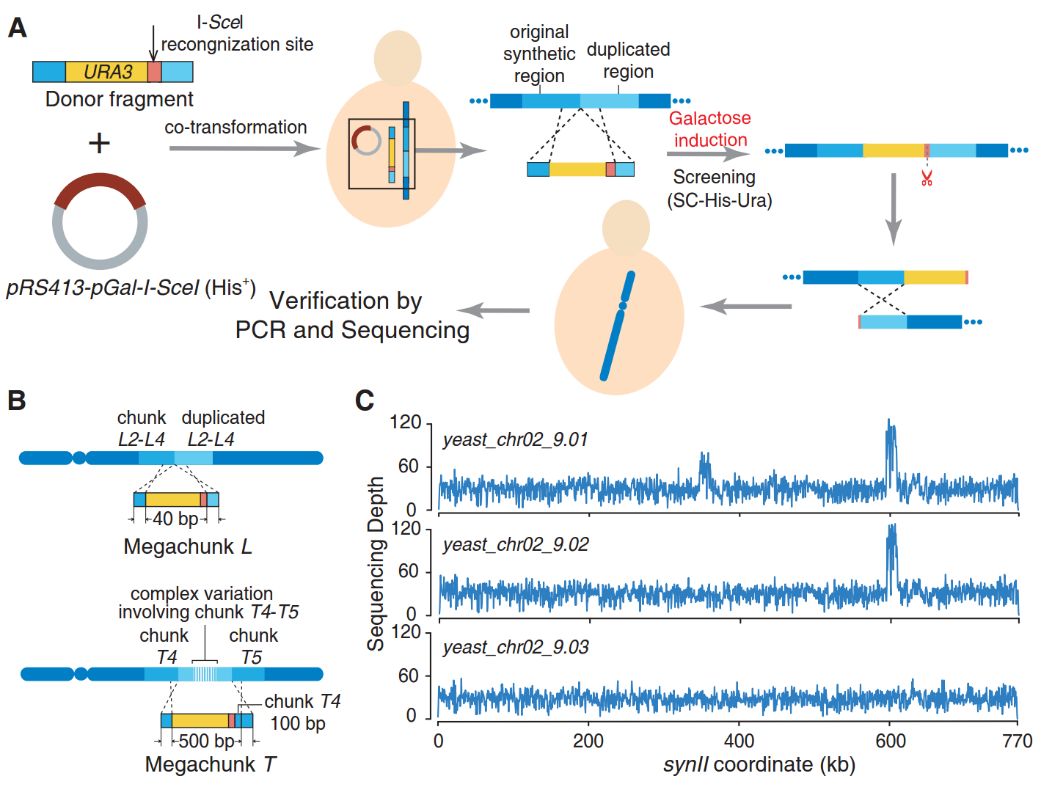
图3 合成染色体结构变异快速修复
合成型酵母菌株表型组特征比较
为了评估人工合成染色体对酿酒酵母细胞生长的影响,研究团队测试了17中不同压力条件下的人工酿酒酵母染色体菌株的生长表型(图 4A),以及富营养培养(YPD)条件下的生长曲线(图4B),表明合成型菌株未见明显的生长相关的表型功能缺陷。
DNA复制与细胞分裂是细胞增值的重要生命过程。为了进一步探究合成染色体对其的影响,研究团队通过GFP荧光标记II号染色体着丝粒,直接观测比较合成型菌株和野生型菌株BY4741的细胞周期过程(图4C),并通过ChIP-seq检测了细胞周期中S时期的合成染色体的DNA复制功能(图4D)。结果表明合成型菌株具有与野生型菌株BY4741较一致的染色体复制和染色体分离过程,对酿酒酵母细胞的增殖与细胞分裂无明显影响。

图4 合成型酵母表型组特征差异比较
合成型酵母菌株贯穿组差异分析
为了在分子层面系统地解析合成菌株与野生型菌株BY4741可能存在的差异,研究团队进行了转录(图5A)、蛋白质(图5B)、代谢(图5C)和脂质 (图5D)多个层次的组学检测与比较分析,发现仅有少数基因或代谢物表现出显著差异,并进一步通过KEGG 和GO功能富集分析,发现与核糖体相关的细胞组成 (核糖体),生物过程 (细胞质翻译) 和分子功能 (核糖体结构组成)均显著上调(图5 E)。由于在合成酵母的基因组中对tRNA进行了删除,故推测核糖体功能上调可能与tRNA缺失有关。为了验证上述猜想,团队在合成酵母菌株中回补了缺失的13个tRNA基因,发现核糖体翻译上调的现象明显缓解(图5F),证明tRNA移除后对于酵母翻译过程的重要影响。综上所述,人工合成酵母II号染色体,在转录组、蛋白组、代谢组和脂质组多个组学层次,表现出与野生染色体相同强健的生物功能,且蛋白质翻译相关功能过程有显著增强。

图5 合成型酵母贯穿组(Trans-Omics)差异比较
研究结论
本研究通过从头设计并构建了一条完整的酿酒酵母II号染色体,带动我国基因组设计、大片段组装以及染色体构建技术取得长足发展。研究团队开发了一种可高效整合并行构建后产生的半合成染色体方法,对未来合成更大规模的染色体基因组意义重大。人工生命的生物学功能表征对于后续合成基因组领域的科研和应用至关重要。研究团队首次整合了生长表型、遗传复制与细胞增殖,以及 “贯穿组学(Trans-Omics)”分析方法,系统性地深度解析了人工合成酵母II号染色体的生物学功能,充分证明了酿酒酵母基因组可增加、可删减的高度灵活性。
综上,合成酿酒酵母II号染色体,是国际合成酿酒酵母基因组计划 (Sc 2.0) 中重要组成部分。开启“设计生命、再造生命和重塑生命”的进程,使我国成为继美国之后第二个掌握真核生物基因组设计构建能力的国家。合成型酿酒酵母菌株可实现在多种压力环境中超速进化,为合成酵母菌株的工业化应用方向注入新的动力,并有望在人类健康、生态、能源、工农业等领域产生重大影响(Yeast 2.0, Nature Communications专辑)。
//
研究数据
本研究涉及组学数据集获取网址:https://www.ncbi.nlm.nih.gov/sra/?term=SRP062892
合成酵母II号染色体序列图谱获取网址:https://www.ncbi.nlm.nih.gov/nuccore/CP013608.1/
